
Page 10 - 《中国药科大学学报》2025年第4期
P. 10
406 学报 Journal of China Pharmaceutical University 2025, 56(4): 405 − 415 第 56 卷
[1]
制,提供快速非特异性防御并激活适应性免疫 。 苷酸合成酶(cyclic GMP–AMP synthase, cGAS)、干
先天免疫系统通过 PRRs 识别 PAMPs 和 DAMPs, 扰素诱导的蛋白 2(absent in melanoma 2, AIM2)、
二者分别是病原微生物所特有的保守成分和损伤 Z-DNA 结合蛋白 1(Z-DNA binding protein 1, ZBP1)
细胞释放的内源性分子 。核酸免疫识别属于天然 等。值得注意的是,核酸识别信号的过度活化或部
[2]
免疫,是先天对病毒感染反应的关键。通常认为宿 分基因缺陷与一系列炎症、自身免疫疾病、癌症等
[4]
主细胞的核酸识别受体能够识别来自病原微生物 疾病密切相关 。因此,核酸免疫识别信号通路
所特有的异源 DNA 或 RNA,感知异常暴露在胞质 需要在分子水平受到严格且精密的调控。本文将
中的自身 DNA,通过多级信号转导,激活下游干扰 论述核酸免疫中不同信号分子(dsDNA、Z-DNA、
素系统、自然杀伤细胞,以达到清除病原体、激活适 RNA)的传感器通路轴(图 1),并简要总结分子机
应性免疫并响应机体损伤的目的 。目前已知的核 制、生物学功能及相关调控分子发现(图 2),聚焦相
[3]
酸 PRRs 主要包括 Toll 样受体 3(Toll like receptors 关疾病及其药物研究,为核酸免疫领域的后续研究
3, TLR3)、Toll 样受体 7-9(Toll like receptors 7-9, 提供新的思路和方向。
TLR7-TLR9)、RNA 传感器视黄酸诱导基因 I 样受
1 RNA 受体介导的免疫反应及其化学干预
体(RIG-I-like receptors, RLRs)、腺苷脱氨酶活性
RNA 1(adenosine deaminase acting on RNA, ADAR1) 病原微生物入侵细胞时,大部分细菌或病毒首
和黑色素瘤分化相关蛋白 5(melanoma differentiation- 先通过细胞的内吞作用进入内体,随后会从内体进
associated gene 5, MDA5)以及环鸟嘌呤-腺嘌呤核 入细胞质中,否则将被运输到溶酶体并降解 。当
[5]
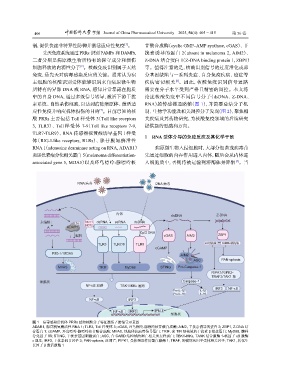
RNA 病毒 DNA 病毒
内体 dsDNA Z-DNA
未编辑 Zα SIDT1 dsRNA ssRNA dsDNA
A ADAR1 SIDT2
CpG DNA ZBP1
I 编辑 cGAS AIM2
TLR3 TLR7/8 TLR9 Zα结构域 RHIMs结构域
cGAMP
RIG-I / MDA5 AIM2
ASC PAN-optosis
MVAS TRIF MyD88 STING Pro-Caspase-1
RIPK1/RIPK3-
TRAF2/TAK1 轴
细胞质 Caspase-1
NF-κB 通路 TBK1/IKKε 通路
Pro-IL-1β IL-1β IRF3 NF-κB
Pro-IL-18 IL-18
NF-κB IRF3
NF-κB IRF3 IFN-I
细胞核
图 1 病毒感染后机体 PRRs 感知核酸分子特征激活下游信号示意图
ADAR1, 腺苷脱氨酶活性 RNA 1; TLR3, Toll 样受体 3; cGAS, 环鸟嘌呤-腺嘌呤核苷酸合成酶; AIM2, 干扰素诱导的蛋白 2; ZBP1, Z-DNA 结
合蛋白 1; cGAMP, 环鸟嘌呤-腺嘌呤核苷酸合成酶; MVAS, 线粒体抗病毒信号蛋白; TRIF, 含 TIR 结构域的干扰素 β 衔接蛋白; MyD88, 髓样
分化因子 88; STING, 干扰素基因刺激蛋白; ASC, 含 CARD 结构域的凋亡相关斑点样蛋白; TBK1-IKKε, TANK 结合激酶 1-核因子 κB 激酶
ε 亚基; IRF3, 干扰素调节因子 3; PAN-optosis, 泛凋亡; PIPK1, 受体相互作用蛋白激酶 1; TRAF, 肿瘤坏死因子受体相关因子; TAK1, 转化生
长因子 β 激活激酶 1