
Page 115 - 《渔业研究》2025年第6期
P. 115
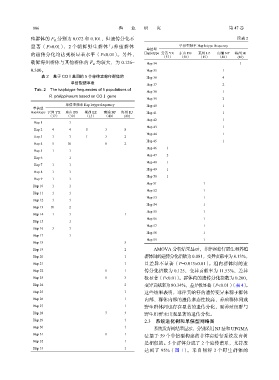
806 渔 业 研 究 第 47 卷
殖群体的 F 分别为 0.072 和 0.101,但遗传分化不 续表 2
t
s
显著(P>0.01) ,2 个朝鲜野生群体与养殖群体 单倍型频率 Hap lotype frequency
单倍型
的遗传分化均达到极显著水平(P<0.01) 。另外, Haplotype 云霄 YX 东山 DS 莱州 LZ 南浦 NP 海州 HJ
(37) (30) (15) (40) (40)
朝鲜海州群体与其他群体的 F 均较大,为 0.126~ Hap 34 1
t
s
0.300。 Hap 35 1
表 2 基于 COⅠ基因的 5 个菲律宾蛤仔群体的 Hap 36 4
单倍型频率表
Hap 37 2
Tab. 2 The haplotype frequencies of 5 populations of
Hap 38 1
R. philippinarum based on COⅠ gene
Hap 39 2
单倍型频率 Hap lotype frequency Hap 40 1
单倍型
Haplotype 云霄 YX 东山 DS 莱州 LZ 南浦 NP 海州 HJ Hap 41 1
(37) (30) (15) (40) (40)
Hap 42 1
Hap 1 1
Hap 43 1
Hap 2 4 4 5 3 8
Hap 44 1
Hap 3 1 1 1 3 2
Hap 45 1
Hap 4 5 10 8 2
Hap 46 1
Hap 5 1 1
Hap 47 2
Hap 6 1
Hap 48 1
Hap 7 1 1
Hap 49 1
Hap 8 1 1
Hap 50 1
Hap 9 1 1
Hap 51 1
Hap 10 1 1
Hap 52 1
Hap 11 1 1
Hap 53 1
Hap 12 1 1
Hap 54 1
Hap 13 10 2
Hap 55 1
Hap 14 1 1 1
Hap 56 1
Hap 15 1
Hap 57 1
Hap 16 3 1
Hap 58 1
Hap 17 1
Hap 59 1
Hap 18 5
Hap 19 2 AMOVA 分析结果显示,菲律宾蛤仔野生和养殖
Hap 20 1 群体间的遗传分化指数为 0.081,变异贡献率为 8.13%,
Hap 21 1 且差异不显著(P=0.013>0.01) 。组内群体间的遗
Hap 22 1 1 传分化指数为 0.125,变异贡献率为 11.53%,差异
Hap 23 1 3 极显著(P<0.01) 。群体内的遗传分化指数为 0.200,
Hap 24 2 变异贡献率为 80.34%,差异极显著(P<0.01)(表 4)。
Hap 25 1 这些结果表明,菲律宾蛤仔的遗传变异来源于群体
Hap 26 1 内部,群体内部的遗传多态性较高,养殖群体间或
Hap 27 1 野生群体间也存在显著的遗传分化,而养殖组群与
Hap 28 7 3 野生组群未出现显著的遗传分化。
Hap 29 1 2.3 系统进化树和单倍型网络图
Hap 30 1 系统发育树结果显示,分别采用 NJ 法和 UPGMA
Hap 31 1 1 法基于 59 个单倍型构建的菲律宾蛤仔系统发育树
Hap 32 1
是相似的。5 个群体分成了 2 个遗传谱系,支持度
Hap 33 1
达到了 95%(图 1) 。来自朝鲜 2 个野生群体的