
Page 63 - 《水产学报》2026年第2期
P. 63
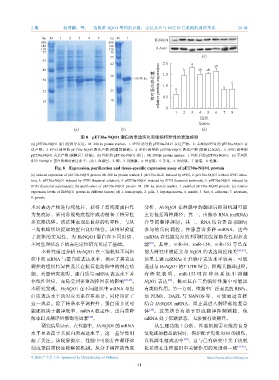
2 期 杨清麟,等:三角帆蚌 NQO1 基因的克隆、表达及其与 Nrf2 信号通路的调控关系 50 卷
ku M 1 2 3 4 5 ku M 1
HcNQO1
200 200
140 140
110 110 β-Actin
75 75
55 (c)
55
42 2.0 a ab
42 b ab ab
相对蛋白表达量 relative protein expression 1.0 c c
30 1.5
30 30.48 ku c
23 23
18 18 0.5
0
1 2 3 4 5 6 7 8
组织
tissues
(a) (b) (d)
图 8 pET30a-NQO1 蛋白的表达纯化和组织特异性的表达检测
(a) pET30a-NQO1 蛋白的诱导表达。M. 200 ku protein marker,1. IPTG 诱导的 pET30a-BL21 表达产物,2. 未添加 IPTG 的 pET30a-NQO1 表
达产物,3. IPTG 诱导的 pET30a-NQO1 表达产物 (细菌裂解液),4. IPTG 诱导的 pET30a-NQO1 表达产物 (裂解后沉淀),5. IPTG 诱导的
pET30a-NQO1 表达产物 (裂解后上清液);(b) 纯化的 pET30a-NQO1 蛋白。M. 200 ku protein marker,1. 纯化后的 pET30a-NQO1;(c) 不同组
织中 HcNQO1 蛋白的相对表达水平;(d) 1. 血淋巴,2. 鳃,3. 肝胰腺,4. 外套膜,5. 斧足,6. 闭壳肌,7. 肠道,8. 性腺。
Fig. 8 Expression, purification and tissue-specific expression assay of pET30a-NQO1 protein
(a) induced expression of pET30a-NQO1 protein. M. 200 ku protein marker, 1. pET30a-BL21 induced by IPTG, 2. pET30a-NQO1 without IPTG induc-
tion, 3. pET30a-NQO1 induced by IPTG (bacterial solution), 4. pET30a-NQO1 induced by IPTG (bacterial sediment), 5. pET30a-NQO1 induced by
IPTG (bacterial supernatant); (b) purification of pET30a-NQO1 protein. M. 200 ku protein marker, 1. purified pET30a-NQO1 protein; (c) relative
expression levels of HcNQO1 protein in different tissues; (d) 1. hemolymph, 2. gills, 3. hepatopancreas, 4. mantle, 5. foot, 6. adductor, 7. intestines,
8. gonads.
术对表达产物进行纯化后,获得了高纯度蛋白作 分析,HcNQO1 在性腺中的翻译后抑制机制可能
为免疫原,采用常规免疫程序成功制备了特异性 主要包括两种路径:其一,由微小 RNA (miRNA)
多克隆抗体。该抗体表现出良好的特异性,与从 介导的翻译抑制;其二,RNA 结合蛋白 (RBPs)
三角帆蚌组织提取的蛋白良好结合。该结果验证 参与的负向调控。性腺富含多种 miRNA,这些
了抗体的实用性,为 HcNQO1 蛋白在不同组织、 miRNA 在性腺发育的不同时期发挥阶段性调控功
不同生理状态下的表达定位研究奠定了基础。 能 [25] 。其中,miR-34、miR-124、miR-153 等已在
本研究通过分析 HcNQO1 在三角帆蚌不同组 前人研究中被证实与 NQO1 的表达调控相关 [26-27] 。
织中的 mRNA 与蛋白质表达水平,揭示了其表达 如果上述 miRNAs 在性腺中表达水平较高,可能
调控的组织特异性及其在抗氧化防御中的潜在功 通过与 HcNQO1 的3' UTR 结合,阻断其翻译过程。
能。大量研究表明,蛋白质与 mRNA 表达水平并 有 研 究 表 明 , miR-153 可 在 神 经 系 统 中 抑 制
非线性对应,而是受到多重调控因素的影响 [22-23] 。 NQO1 表达 [28] ,提示其在三角帆蚌性腺中可能具
本研究发现,HcNQO1 在不同组织中 mRNA 与蛋 有类似作用。另一方面,性腺中广泛表达的 RBPs,
白质表达水平的对应关系存在差异,同样印证了 如 PUM1、DAZL 与 NANOS 等,可能通过直接
这一观点。除了转录水平调控外,蛋白质丰度可 结合 HcNQO1 mRNA,阻止其进入翻译起始复合
能还取决于翻译效率、mRNA 稳定性、蛋白质降 体 [29] 。这类结合有助于形成翻译抑制颗粒,使
解率以及翻译后修饰等因素 。 mRNA 处于沉默状态,无法被有效翻译。
[24]
研究结果显示,在性腺中,HcNQO1 的 mRNA 从生理功能上分析,性腺细胞需在维持自身
水平显著高于其蛋白质表达水平,这一差异性引 氧化还原稳态的同时,保护配子免受 ROS 的损伤,
[30]
起了关注。该现象提示,性腺中可能存在翻译抑 以保障生殖成功率 ,这与已有研究中关于抗氧
制或蛋白质快速降解的机制。从分子调控的角度 化系统在生殖组织中关键作用的发现相一致 [31-32] 。
中国水产学会主办 sponsored by China Society of Fisheries https://www.china-fishery.cn
11