
Page 148 - 《水产学报》2023年第1期
P. 148
李晓晖,等 水产学报, 2023, 47(1): 019610
5 4 c 1 200 b
1 100
HIF-1α 相对表达量 relative expression of HIF-1α 3 2 a b ab HIF-1α 相对表达量 relative expression of HIF-1α 100 c ac
1 000
80
60
1.5
0.5
0 1 1.0 0 a
1 2 3 4 1 2 3 4
不同低氧胁迫组 不同低氧胁迫组
different hypoxic stresses groups different hypoxic stresses groups
(a) (b)
260 b 20 b
180
HIF-1α 相对表达量 relative expression of HIF-1α 100 a c d HIF-1α 相对表达量 relative expression of HIF-1α 15 5 c d
20
10
1.5
1.0
0.5
0 0 a
1 2 3 4 1 2 3 4
不同低氧胁迫组 不同低氧胁迫组
different hypoxic stresses groups different hypoxic stresses groups
(c) (d)
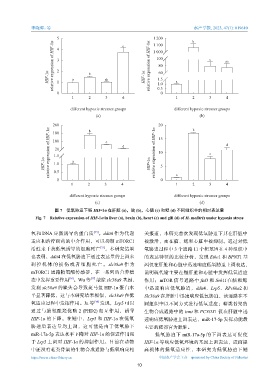
图 7 低氧胁迫下鲢 HIF-1α 在肝脏 (a),脑 (b),心脏 (c) 和鳃 (d) 不同组织中的相对表达量
Fig. 7 Relative expression of HIF-1α in liver (a), brain (b), heart (c) and gill (d) of H. molitrix under hypoxia stress
氧和 DNA 应激诱导的蛋白质 [35] 。ddit4 作为代谢 关报道,本研究首次发现低氧胁迫下其在肝脏中
适应和治疗耐药的中介作用,可以抑制 mTORC1 被激活,而在脑、鳃和心脏中被抑制。通过对低
活性来干扰低氧诱导的细胞死亡 ,本研究结果 氧胁迫过程中 3 个通路 11 个靶基因在 4 种组织中
[22]
也表明,ddit4 在低氧胁迫下通过表达量的上调来 的表达特征的比较分析,发现 Ethe1 和 BPNT1 基
调 控 机 体 的 损 伤 或 者 细 胞 死 亡 。 slc38a9 作 为 因仅在肝脏和心脏中迅速响应低氧胁迫上调表达,
mTORC1 通路精氨酸传感器,在一系列的营养稳 说明硫代谢主要在鲢肝脏和心脏中发挥低氧适应
[36]
态中发挥重要作用 。Wu 等 [37] 敲除 slc38a9 基因, 作用;mTOR 信号通路中 fzd3 和 Seh1l 在脑和鳃
发现 slc38a9 的缺失会导致斑马鱼 HIF-1α 蛋白水 中迅速响应低氧胁迫,ddit4、Lrp5、RPs6ka2 和
平显著降低,这与本研究结果相似,slc38a9 在低 Slc38a9 在肝脏中迅速响应低氧胁迫,该通路在不
氧适应过程中发挥作用。Ju 等 [38] 发现,Lrp5 可以 同组织中以不同方式进行低氧适应;萜类骨架的
通过与脯氨酰羟化酶 2 (PHD2) 相互作用,诱导 生物合成通路中的 icmt 和 PCYOX1 仅在肝脏中迅
HIF-1α 的下降。在鲢中,Lrp5 和 HIF-1α 在低氧 速响应低氧胁迫上调表达,miR-17-5p 发挥功能最
胁迫后表达量均上调,这可能是由于低氧胁下 主要的靶器官为肝脏。
miR-17a-5p 表达水平下降对 HIF-1α 的促进作用强 低氧胁迫下 miR-17a-5p 的下调表达可促使
于 Lrp5 上调对 HIF-1α 的抑制作用。目前在动物 HIF-1α 等响应低氧环境的基因上调表达,进而提
中还没有萜类骨架的生物合成通路与低氧响应相 高机体的低氧适应性,本研究为低氧胁迫下鲢
https://www.china-fishery.cn 中国水产学会主办 sponsored by China Society of Fisheries
10