
Page 124 - 《渔业研究》2026年第1期
P. 124
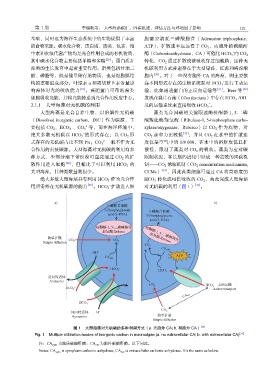
第 1 期 李瑞帆等: 大型海藻碳汇:固碳机理、评估方法与环境因子影响 121
基础,同时也为海洋生态系统中的生物提供了丰富 胞需要消耗三磷酸腺苷(Adenosine triphosphate,
的食物来源。碳水化合物、蛋白质、脂类、色素、维 ATP) ,扩散速率远远慢于 CO ,而胞外的碳酸酐
2
生素和次级代谢产物均是光合作用合成的有机物质, 酶(Carbonicanhyd-rase,CA)可催化 HCO 向 3 − CO 2
其中碳水化合物主要包括单糖和多糖 [26] ;蛋白质在 转化,CO 通过扩散或被吸收穿过细胞膜,这种无
2
藻类的生长发育中起着重要作用;脂类包括甘油三 机碳利用方式普遍存在于大型绿藻、红藻和褐藻细
酯、磷脂等,既是能量储存的物质,也是细胞膜结 胞内 [30] 。对于一些没有胞外 CA 的海藻,则主要依
构的重要组成部分;叶绿素 a 和类胡萝卜素含量影 靠不同形式存在的生物系统泵对 HCO 进行主动运
−
3
响藻体对光的吸收能力 [27] ;藻胆蛋白可帮助藻类 输,比如通透蛋白的正反向运输等 [31] 。Beer 等 [32]
细胞吸收光能,并将光能传递到光合作用反应中心。 发现在裂片石莼(Ulva fasciata)中存在 HCO /OH −
−
3
2.1.1 大型海藻对无机碳的利用 反向运输系统来直接吸收 HCO 。
−
3
大型海藻是光合自养生物,以溶解性无机碳 藻类光合固碳的关键限速酶核酮糖 1, 5-二磷
(Dissolved inorganic carbon,DIC)作为碳源,主 酸羧化酶/加氧酶(Ribulose-1, 5-bisphosphate carbo-
要包括 CO 、HCO 、CO 等,而在海洋环境中, xylase/oxygenase,Rubisco)以 CO 作为底物,对
2−
−
2
2
3
3
绝大多数无机碳以 HCO 的形式存在;以 CO 形 CO 亲和力比较低 [33] ,并且 CO 在水中的扩散速
−
2
2
2
3
2−
式存在的无机碳占比不到 1%;CO 一般不作为光 度仅是空气中的 1/8 000,在水中的溶解度低且扩
3
合作用的直接碳源。大型海藻对无机碳的利用有多 散慢,限制了藻类对 CO 的吸收。藻类为应对碳
2
种方式,少部分潮下带红藻可直接通过 CO 的扩 限制状况,在长期的进化中形成一种高效的固碳机
2
−
散作用进入细胞 [28] ,但相比于可以利用 HCO 的 制——CO 浓缩机制(CO concentration mechanism,
2
2
3
大型海藻,其种类数量则很少。 CCMs) [34] ,因此藻类细胞可通过 CA 将高浓度的
绝大多数大型海藻具有利用 HCO 作为光合作 HCO 转化成可供吸收的 CO ,由此完成大型海藻
−
−
3
3
2
−
用所需外在无机碳源的能力 [29] 。HCO 扩散进入细 对无机碳的利用(图 1) [35] 。
3
a) b)
3-磷酸甘油酸
3-Phosphoglyceric 3-磷酸甘油酸
acid (3-PGA) 3-Phosphoglyceric
acid (3-PGA)
核酮糖-1, 5-二磷酸羧化
加氧酶 Rubisco 核酮糖-1, 5-二磷酸羧化
简单扩散 加氧酶 Rubisco
CO 2 CO 2
Simple diffusion
CO 2
CA cyto
OH −
ATP
CA cyto
−
HCO 3 −
HCO 3
逆向转运体
Antiporter
− 主动运输
CO 2 HCO 3
−
HCO 3 Active transport
CA ext
−
HCO 3
同向转运体 H + CO 2
Symporter 简单扩散
Simple diffusion
图 1 大型海藻对无机碳的多种利用方式(a. 无胞外 CA;b. 有胞外 CA) [35]
Fig. 1 Multiple utilization modes of inorganic carbon in macroalgae (a. no extracellular CA; b. with extracellular CA) [35]
注:CA cyto 为胞质碳酸酐酶;CA ext 为胞外碳酸酐酶。以下同此。
Notes: CA cyto is cytoplasm carbonic anhydrase; CA ext is extracellular carbonic anhydrase. It’s the same as below.