
Page 82 - 《水产学报》2026年第04期
P. 82
4 期 水 产 学 报 50 卷
N 2010 N 2011 N 2012 N 2013 N 2014
36° 36° 36° 36° 36°
33° 33° 33° 33° 33°
30° 30° 30° 30° 30°
27° 27° 27° 27° 27°
120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E
(a) (b) (c) (d) (e)
N 2015 N 2016 N 2017 N 2018 N 2019
36° 36° 36° 36° 36°
33° 33° 33° 33° 33°
30° 30° 30° 30° 30°
27° 27° 27° 27° 27°
120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E
(f) (g) (h) (i) (j)
N 2020 N 2022 N 2023
36° 36° 36°
ln[CPUE/(g/h)]
33° 33° 33°
4
30° 30° 30° 2
0
27° 27° 27° −2
120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E 120° 122° 124° 126° 128°E
(k) (l) (m)
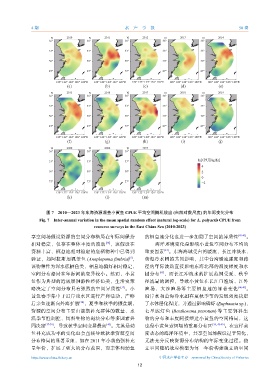
图 7 2010—2023 年东海资源调查小黄鱼 CPUE 平均空间随机效应 (自然对数尺度) 的年际变化分布
Fig. 7 Inter-annual variation in the mean spatial random effect (natural log-scale) for L. polyactis CPUE from
resource surveys in the East China Sea (2010-2023)
享空间场假设资源的空间分布格局在年际间保持 的栖息地分化也进一步加剧了空间的异质性 [41-43] 。
[36]
相对稳定,仅存在整体丰度的波动 ,该假设在 海洋环境变化是影响小黄鱼空间分布不均的
资源丰富、栖息地相对稳定的底栖物种中已得到 重要因素 。东海海域受台湾暖流、长江冲淡水、
[44]
验证,如阿拉斯加裸盖鱼 (Anoplopoma fimbria) , 黄海冷水团的共同影响,其中台湾暖流强度和路
[5]
该物种作为深水底栖鱼类,栖息地偏好相对稳定, 径的年际波动直接影响东海北部的温度梯度和水
[45]
空间分布格局在年际间的变异较小。然而,小黄 团分布 ,而长江冲淡水的扩展范围受夏、秋季
鱼作为典型的近底层洄游性经济鱼类,生活史策 径流量的调控,导致小黄鱼在长江口渔场、江外
[37]
略决定了空间分布具有强烈的空间异质性 。小 渔场、大沙渔场等主要栖息地的显著变化 [46-47] 。
黄鱼春季集中于近岸浅水区进行产卵活动,产卵 沿岸水和黄海冷水团在夏秋季节的发展强度决定
后亲鱼逐渐向外海扩散 ,夏季和秋季的摄食期, 了水体层化程度,并通过影响磷虾 (Euphausia sp.)、
[38]
资源的空间分布主要由前期补充群体的数量、水 七星底灯鱼 (Benthosema pterotum) 等主要饵料生
温季节性调控、饵料生物的斑块分布等多因素共 物的分布和丰度间接塑造小黄鱼的空间格局,这
[40]
同决定 [37-39] ,导致秋季空间变异最高 。尤其是幼 也是小黄鱼索饵场的重要分布区 [39, 48-49] 。在这样高
鱼补充成功率的变化也会直接导致秋季资源空间 度动态的海洋环境中,共享空间场假设过于简化,
分布格局的显著重组,如在 2011 年小黄鱼强补充 无法充分反映资源分布结构的年际变化过程。独
量年份,扩展了更大的分布范围,而亲体和幼鱼 立空间随机效应模型为每一年份构建独立的空间
https://www.china-fishery.cn 中国水产学会主办 sponsored by China Society of Fisheries
12